La reproduction sexuée des plantes à fleur
La reproduction sexuée, c’est-à-dire la formation d’un nouvel individu à partir d’un œuf issu de la fusion de cellules reproductrices femelles et mâles, existe chez toutes les plantes. Ces dernières sont d’ailleurs qualifiées d’embryophytes (embryo = embryon ; -phyte = plante) pour traduire le fait que toute nouvelle plante issue de la reproduction sexuée résulte du développement d’un embryon. Toutefois, les modalités de la reproduction sexuée sont différentes chez les différents groupes de plantes, ce qui reflète l’histoire évolutive de ces êtres vivants. On ne considère ici que la reproduction sexuée des plantes à fleur.
Les plantes à fleur, ou spermatophytes (spermato = semence, graine ; -phyte = plante), sont caractérisées, comme leur nom l’indique, par la formation de graines à l’issue de la reproduction sexuée. Contrairement aux mousses et aux fougères, la dissémination de ces plantes ne se fait pas par des spores mais par les graines. La graine n’existe dans aucun autre groupe végétal et son « invention » a constitué une innovation évolutive majeure, à mettre en relation avec le succès de ces plantes au cours des temps géologiques.
Les spermatophytes regroupent l’ensemble des arbres, arbustes et plantes herbacées à fleur. On les divise en gymnospermes et angiospermes.
Les gymnospermes (gymno = nu ; sperme = semence, graine) sont des plantes ligneuses vivaces (arbres, arbustes et lianes) caractérisées par des graines nues, c’est-à-dire non incluses dans un fruit.
Les angiospermes (angio = capsule ; sperme = semence, graine) sont les plantes à fleur proprement dites et sont caractérisées par leurs ovules enclos dans un organe spécialisé appelé carpelle. L'ensemble des carpelles est appelé ovaire. À la suite de la fécondation, les ovules se transforment en graines et sont enfermées dans un fruit résultant de la transformation de l'ovaire.
Chez les gymnospermes, comme par exemple les conifères, les organes reproducteurs sont des « fleurs » simples, appelées cônes. Les cônes femelles, de petite taille, sont constitués d’un axe portant des écailles auxquelles sont fixés deux ovules nus, c’est-à-dire qui ne sont pas inclus dans un carpelle, contrairement à ceux des angiospermes. Les cônes mâles présentent une organisation similaire avec un axe portant des écailles. Chaque écaille comporte deux sacs polliniques dans lesquels se forme le pollen et correspondent donc aux étamines des fleurs d’angiospermes. À maturité, le pollen s’échappe des sacs polliniques et la présence d’une paire de ballonnets sur chaque grain de pollen facilite son transport par le vent.

Cônes mâles et femelles (Chamaecyparis ou faux cyprès)
La fleur des angiospermes
Si les fleurs des angiospermes sont, le plus souvent, hermaphrodites, c’est-à-dire comportant à la fois des organes mâles et des organes femelles, il existe aussi des espèces de plantes à fleurs dont les sexes sont séparés, certains plants portant des fleurs mâles, d’autres des fleurs femelles.
Contrairement aux fleurs simples des conifères et autres gymnospermes, la fleur des angiospermes comporte généralement quatre types de pièces florales différentes, deux types de pièces stériles, les sépales et les pétales, entourant deux types de pièces fertiles, mâles et femelles, appelées respectivement étamines et pistil, qui produisent les cellules reproductrices. Le nombre, la position et la forme des différentes pièces florales sont spécifiques de chaque espèce de plante à fleurs et sont utilisés dans les clés de détermination permettant de les identifier.

Structure d'une fleur (cerisier)
Certaines espèces de plantes présentent des fleurs auxquelles certaines pièces manquent. On parle alors de fleurs incomplètes. C’est, par exemple, le cas des fleurs du chêne ou du hêtre qui sont dépourvues de pétales.
Chez de nombreuses plantes, les fleurs constituent une inflorescence, c’est-à-dire que plusieurs fleurs sont regroupées, de façon plus ou moins dense, à l’extrémité d’un même pédoncule qui porte alors le nom de hampe florale. C’est le cas, par exemple, des marguerites ou des tournesols. Contrairement à ce qu’indique le langage courant, ce qui est appelé « fleur » chez ces plantes, correspond, en réalité, à de nombreuses petites fleurs serrées sur un même réceptacle floral. Comme chaque fleur périphérique comporte un seul pétale, tandis que les fleurs centrales en sont dépourvues, l’inflorescence donne l’impression d’être une fleur simple.

Inflorescences (pâquerette)
La pollinisation
La pollinisation est le processus par lequel les grains de pollen sont transportés d’une fleur à une autre, permettant ainsi le rapprochement des gamètes et la fécondation. Les mécanismes de la pollinisation diffèrent selon les espèces de plantes. Les agents de la pollinisation sont le vent et les animaux, en particulier les insectes.
Ils sont attirés notamment par les couleurs des fleurs, leurs odeurs ou le nectar, un liquide sucré produit par des glandes spécialisées de certaines fleurs. Lorsque les insectes butinent les fleurs, le pollen s'accroche à leur corps et peut être ainsi transporté sur d'autres fleurs. Quelque 100 000 espèces animales différentes participent ainsi à la reproduction sexuée des plantes à fleurs et plus des deux tiers des plantes cultivées dépendent d’animaux pour leur pollinisation.

Abeille butineuse
Noter la boulette de grains de pollen agglomérés sur la patte postérieure
La fécondation
La fécondation est la formation d’un œuf résultant de la fusion des cellules reproductrices mâle et femelle. La fécondation se produit à la suite de la pollinisation, quand des grains de pollen ont été déposés sur le stigmate, partie terminale du pistil. Les grains de pollen adhèrent au stigmate en raison des papilles gluantes qui le recouvrent. Une fois collé au stigmate, le grain de pollen germe en produisant un tube, le tube pollinique, qui s’allonge à travers les tissus du pistil jusqu’à atteindre l’ovaire contenant les ovules. En raison de l’existence du tube pollinique, la fécondation des plantes à graine est indépendante du milieu aquatique, contrairement à celle des mousses et des fougères, par exemple.
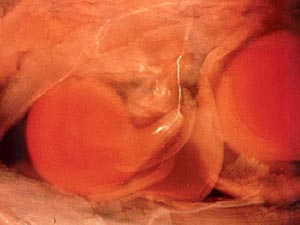
Fécondation du colza. Un tube pollinique (révélé par épifluorescence, en blanc) véhicule les gamètes mâles jusqu'à l'ovule (en rouge)
Fruits et légumes
Dans le vocabulaire de la botanique, les termes de fruit et légume ont une signification précise. On appelle fruit l’organe contenant les graines qui résulte de la transformation de l’ovaire de la fleur à la suite de la fécondation. On appelle légume ou gousse le fruit sec déhiscent (c'est à dire qui s'ouvre à maturité pour libérer ses graines) de certaines espèces de plantes, en particulier celui des fabacées (anciennement appelées légumineuses), famille à laquelle appartiennent les fèves, les haricots et les pois.

Fleurs et fruits (gousse) du genêt
La gousse est un légume au sens botanique
Il n’en est pas de même dans le langage courant. Dans le domaine alimentaire, on parle de légume pour désigner indistinctement divers organes végétaux qui se consomment généralement salés en garniture. Il peut s’agir de tiges (poireau, fenouil), de feuilles (épinards, choux, salades), de fruits (tomate, haricots verts, courgette, aubergine), de graines (pois, haricots blancs) de racines (carottes), de bourgeons (asperges), d’inflorescences (chou-fleur), etc., alors que le terme de fruit du langage courant recouvre simplement les fruits à goût sucré, généralement consommés en dessert. Mais tous les fruits, au sens botanique, ne sont pas nécessairement sucrés, comme les haricots verts, les courgettes, les aubergines, etc., et la signification botanique des termes de fruit et de légume ne doit pas être assimilée à celle du langage courant.
La graine
La graine, organe caractéristique de la reproduction sexuée des plantes à fleur, est le résultat de la double fécondation d’un ovule par un grain de pollen produisant, d’une part, une cellule-œuf à l’origine d’un embryon et, d’autre part, un tissu de réserve. Généralement libérées à la fin de l’été, les graines permettent à la fois la dissémination à distance des plantes et le passage de la mauvaise saison.
Typiquement, toutes les graines comportent un embryon et des réserves, le tout étant enveloppé par un ou plusieurs téguments.
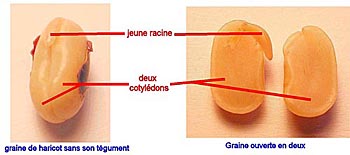
Structure de la graine de haricot
Un embryon est constitué d’une plantule portant un, deux ou plusieurs cotylédons, selon les groupes de plantes à fleur. La plantule est elle-même formée d’une radicule, à l’origine de la racine, d’une tigelle, à l’origine de la tige, sur laquelle sont fixés le ou les cotylédons et d’un bourgeon terminal, qualifié de gemmule. Les cotylédons sont des feuilles particulières qui ne se forment que dans les graines.

Détails de la plantule de la graine de haricot
Les plantes dont les graines ne comportent qu’un seul cotylédon sont qualifiées de monocotylédones et sont également caractérisées par les nervures parallèles de leurs feuilles, comme chez les graminées (blé, maïs, riz) ou les liliacées (lis, tulipe).

Parterre d'iris
Les iris appartiennent aux monocotylédones. Les nervures de leurs feuilles sont parallèles
Celles dont les graines comportent deux cotylédons sont qualifiées de dicotylédones et sont également caractérisées par les nervures ramifiées de leurs feuilles. C’est dans ce groupe que l’on trouve la plus grande diversité d’espèces.

Pissenlits, lamiers blancs, bouton d'or, etc.
Les plantes à fleur les plus nombreuses sont des dicotylédones. Les nervures de leurs feuilles sont ramifiées
Enfin, chez les gymnospermes, comme les conifères, la graine comporte un nombre de cotylédons supérieur à deux.
En climat tempéré, les graines sont capables de passer l’hiver en résistant au froid, à la sécheresse ou à l’excès d’humidité. La résistance des graines est liée essentiellement à leur déshydratation poussée qui leur permet de se maintenir en vie ralentie et à leurs téguments qui les empêche de se réhydrater prématurément.
Semence, grain et graine
Ce n’est pas toujours la graine proprement dite qui, une fois semée, est susceptible de donner naissance à une nouvelle plante. En effet, chez certaines plantes, comme les graminées (blé, maïs, riz), le grain correspond en réalité à un fruit sec (akène) dont l’enveloppe est soudée aux téguments de la graine, ce qui les rend difficile à distinguer.
De façon générale, on appelle semence le grain qui, une fois semé, est capable de donner naissance à une nouvelle plante, qu’il corresponde à une graine ou à un fruit. Ainsi, les semences de haricot, de lentille, de fève, de melon, de citrouille, de pommier, etc. sont des graines alors que les semences de graminées (maïs, riz, blé) sont des fruits secs qualifiés de caryopses. Il faut noter cependant que les grains de riz du commerce alimentaire sont des caryopses débarrassés de leurs enveloppes (qui constituent le son), mais ils sont incapables de germer ainsi (riz poli). Seuls les grains de riz non décortiqués (riz complet) sont susceptibles de germer.
La reproduction asexuée ou multiplication végétative
La multiplication végétative est une forme de reproduction dans laquelle une partie de l’appareil végétatif (la partie non reproductrice de la plante), racine, tige ou feuille, est susceptible de redonner naissance à une plante entière. Comme la multiplication végétative ne fait intervenir aucun phénomène sexuel, la plante obtenue est génétiquement identique à la plante mère. On appelle clone un organisme génétiquement identique à celui qui lui a donné naissance. La capacité des végétaux à se reproduire de façon asexuée par multiplication végétative est liée à la grande plasticité des tissus végétaux. Les cellules végétales elles-mêmes sont capables de se dédifférencier avant de se multiplier et de donner un massif de cellules indifférenciées, appelé cal, susceptible de redonner naissance à une plante complète.

La multiplication végétative permet à certaines plantes, ici la joubarbe, d'envahir leur substrat
La multiplication végétative naturelle se produit à partir des organes les plus divers. Il peut s’agir d’organes souterrains, comme les rhizomes (iris, chiendent), les bulbes (tulipe, ail, poireau), les tubercules (pomme de terre), les racines (lilas, framboisier).

Formation de nouvelles tiges par multiplication végétative à partir d'un tubercule de pomme de terre
Dans ce dernier cas, on parle de drageons. Il peut aussi s’agir d’organes aériens, comme les tiges, les feuilles, voire les fleurs. Ainsi, chez le fraisier, la plante donne naissance à un axe horizontal, appelé stolon ou coulant, sur lequel se développent de nouveaux pieds qui restent ensuite séparés lorsque le stolon disparaît. Chez la ronce, lorsqu’un rameau aérien touche le sol, des racines puis une tige se développent et donnent naissance à un nouveau pied.
La multiplication végétative artificielle est exploitée, non seulement en agriculture, mais aussi en laboratoire, pour multiplier et propager les plantes alimentaires ou ornementales.
Le bouturage consiste à reproduire une plante en isolant un organe ou un fragment d’organe que l’on place dans l’eau ou en terre. Une fois des racines formées, il se développe un nouvel individu. Des préparations à base d’hormones végétales peuvent être utilisées pour faciliter le bouturage. Le bouturage artificiel peut-être réalisé à partir de jeunes tiges chez un grand nombre d’espèces (géranium, osier, rosier, saule, groseillier, laurier-rose, forsythia, yucca, etc.) ou de feuilles (bégonia, Saintpaulia). Toutefois, certaines espèces sont impossibles à bouturer.
Le marcottage peut être forcé chez certaines espèces, comme la vigne ou l’œillet en enterrant une tige encore portée par la plante. Lorsque des racines, puis une nouvelle tige se développent, on sépare le nouveau pied de la plante mère.
La greffe est surtout utilisée pour multiplier des arbres fruitiers. Elle consiste à insérer une tige ou un bourgeon (greffon) sur un arbre ou arbuste d’une espèce proche ou d'une variété plus robuste (porte-greffe) de façon à mettre en contact étroit les tissus des deux plantes jusqu’à ce que des connexions s’établissent entre leurs vaisseaux conducteurs. Cette méthode permet de multiplier une plante d’intérêt, par exemple produisant de gros fruits, en profitant des qualités du porte-greffe, comme la vigueur ou la résistance. On multiplie ainsi certains poiriers en greffant leurs branches sur un tronc de cognassier.
Outre ces méthodes, connues depuis des temps très anciens, il est possible aujourd’hui de multiplier les plantes, quasiment à l’infini, par microbouturage et culture in vitro qui permettent la multiplication en grande quantité de plantes à partir de quelques cellules isolées d’une plante mère. Ainsi, la mise en culture in vitro d’explants isolés à partir d’une seule feuille de Saintpaulia permet de produire en un an plus de 3 millions de plantes identiques, là où le bouturage classique ne permet d’en obtenir que 675.